Impact Factor ISSN: 1449-1907

 Global reach, higher impact
Global reach, higher impactInt J Med Sci 2017; 14(10):984-993. doi:10.7150/ijms.20336 This issue Cite
Research Paper
Kaempferol Inhibits the Invasion and Migration of Renal Cancer Cells through the Downregulation of AKT and FAK Pathways
Tung-Wei Hung1, 2, Pei-Ni Chen3, 4, Hsu-Chen Wu5, Sheng-Wen Wu2, Pao-Yu Tsai2, Yih-Shou Hsieh3, 4 ![]() , Horng-Rong Chang2, 6
, Horng-Rong Chang2, 6 ![]()
1. Institute of Medicine, Chung Shan Medical University, Taichung, Taiwan;
2. Division of Nephrology, Department of Internal Medicine, Chung Shan Medical University Hospital, Taichung, Taiwan;
3. Clinical Laboratory, Chung Shan Medical University Hospital, Taichung, Taiwan;
4. Institute of Biochemistry and Biotechnology, Chung Shan Medical University, Taichung, Taiwan;
5. Division of Nephrology, Department of Internal Medicine, Erlin Branch of Changhua Christian Hospital, Changhua County, Taiwan;
6. School of Medicine, Chung Shan Medical University, Taichung, Taiwan.
Received 2017-3-30; Accepted 2017-6-18; Published 2017-8-18
Abstract
Kaempferol, which is isolated from several natural plants, is a polyphenol belonging to the subgroup of flavonoids. Kaempferol exhibits various pharmacological activities, including anti-inflammatory, antioxidant, antimicrobial, and anticancer activities. In this study, kaempferol can significantly inhibit the invasion and migration of 786-O renal cell carcinoma (RCC) without cytotoxicity. We examined the potential mechanisms underlying its anti-invasive activities on 786-O RCC cells. Western blot was performed, and the results showed that kaempferol attenuates the manifestation of metalloproteinase-2 (MMP-2) protein and activity. The inhibitive effect of kaempferol on MMP-2 may be attributed to the downregulation of phosphorylation of Akt and focal adhesion kinase (FAK). By examining the SCID mice model, we found that kaempferol can safely inhibit the metastasis of the 786-O RCC cells into the lungs by about 87.4% as compared to vehicle treated control animals. In addition, the lung tumor masses of mice pretreated with 2-10 mg/kg kaempferol were reduced about twofold to fourfold. These data suggested that kaempferol can play a promising role in tumor prevention and cancer metastasis inhibition.
Keywords: renal cell carcinoma, kaempferol, metastasis, invasion.
Introduction
Renal cell carcinoma (RCC) originates from renal parenchyma and is the most common primary renal tumor. It accounts for more than 90% of the cases. RCC incidence rates vary from region to region. In particular, RCC incidence rates in Europe and North America are high, whereas those Asia and South America are low. Furthermore, RCC incidence rate can be as high as 15.3 per 100,000 population (Czech Republic) to 2.8-5.8 per 100,000 population (Asian country). Men are twofold more prone to develop kidney cancer than women worldwide [1]. According to the National Cancer Institute (NCI) Surveillance, Epidemiology, and End Results (SEER) registry (2003-2007), RCC occurs predominantly in the sixth to eighth decade of life, with a median age at diagnosis of around 64 years, and more than 16% of patients with kidney cancer present metastatic diseases at diagnosis because of their silent clinical course [2-4]. Surgical treatment is the main treatment for localized kidney tumor, while medical therapies are commonly suggested for advanced RCC. However, unresectable or metastatic RCC generally presents poor prognosis. The 5-year survival rate in RCC patients with tumour, node and metastasis (TNM) stage IV is approximately 20% [5]. Poor prognosis may be related to the inert response of metastatic RCC to medical therapy.
Metastasis, which involves several processes, including extracellular matrix (ECM) degradation and remodeling, is the leading cause of cancer death [6]. Numerous proteinases involved in the interaction between ECM and cancer cells include matrix metalloproteinases (MMPs), tissue inhibitor metalloproteinase proteins (TIMPs), and urokinase plasminogen activator. Considerable efforts have been devoted to elucidate the mechanisms underlying tumor invasion and metastasis, in which gelatinases (MMP-2 and MMP-9) are vital factors [7]. Several studies indicated that MMP-2 and MMP-9 overexpression can correlate to cancer metastasis in oral cancer [8], breast cancer [9], lung cancer [10], prostate cancer [11], and kidney cancer [12]. Moreover, MMP-2 level is considered as a predictor of tumor recurrence and prognosis [13].
Kaempferol is a natural phytochemical belonging to the flavonoid group. As indicated by several documented works, these polyphenolic compounds, especially kaempferol, possess anticancer properties apart from their antioxidative effects [14, 15]. Previous studies showed that kaempferol can inhibit cancer cell invasion and migration in various human cancer cell lines, such as nonsmall cell lung cancer [16], breast cancer [17], oral cavity cancer [18], medulloblastoma [19], colon cancer [20], and leukemia [21]. Furthermore, kaempferol exerts an apoptotic effect by inactivating Akt in human leukemia cells [22], whereas caspase-dependent apoptosis was noted in oral cavity cancer cells [23]. Apart from its extensive pharmacological benefits, kaempferol is well tolerated and mostly free of toxicity. Thus, the present study investigated the molecular mechanisms underlying the effect of kaempferol on the invasion and migration of RCC 786-O cell in vitro and in vivo.
Materials and Methods
Cell Culture and Kaempferol Treatment
786-O (a human renal cell adenocarcinoma) and human kidney-2 (HK-2; a human proximal tubule epithelial cell line) cells were obtained from the Bioresource Collection and Research Center (Hsinchu, Taiwan). The 786-O cells were cultured in an RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS), 2 mM of L-glutamine, 100 units/mL of penicillin, and 100 μg/mL of streptomycin; HK-2 was cultured in 1:1 mixture of DMEM and Ham's F12 medium containing 10% fetal bovine serum. All the cell cultures were maintained at 37 °C in a humidified 5% CO2 atmosphere. For kaempferol treatment, adequate amounts of stock solution (0.1 M in dimethyl sulfoxide) of kaempferol were added into the culture medium to achieve the indicated concentrations. The resulting solutions were then incubated with the cells at indicated time periods. Dimethyl sulfoxide solution without kaempferol was used as blank reagent.
Determination of Cell Viability (Microculture tetrazolium (MTT) Assay)
To determine the cytotoxicity of kaempferol, we performed a microculture tetrazolium (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide) colorimetric assay to evaluate cell viability. 786-O and HK-2 cells were seeded in 24-well plates at a density of 3 × 104 cells/well and treated with kaempferol at concentrations ranging from 0 μM to 100 μM at 37 °C for 24 h. After the exposure period, the media were removed, and the cells were washed with phosphate buffered saline and incubated with MTT (5 mg/mL) for 4 h. The viable cell number per dish is directly proportional to the production of formazan and can be measured by a Hitachi U-1900 spectrophotometer at 563 nm after the solubilization with isopropanol [12].
Cell Migration and Invasion Assays
First, RCC 786-O cells were treated with different concentrations of kaempferol (0, 25, 50, 75, and 100 μM) for 24 h. The surviving cells were harvested and seeded in a Boyden chamber (Neuro Probe, Cabin John, MD, USA) at 1.5×104/well in a serum-free medium and then incubated for 6 h at 37 °C. For the invasion assay, 100 μg/cm2 of Matrigel (25 mg/50 mL; BD Biosciences, MA, USA) was applied to the cellulose nitrate filters with a standard medium covering the bottom chamber. Filters were then air-dried for 5 h in a laminar flow. The invaded cells were fixed with 100% methanol and stained with 5% Giemsa. Cell numbers were counted under a light microscope (×100). Migration assay was performed as described in the invasion assay without Matrigel coating [12].
Wound healing assay
We determined whether kaempferol could alter the migration of 786-O cells. A wound closure seeding model was constructed using silicon culture inserts (Ibidi, Munchen, Germany) with two individual wells for cell seeding. Each insert was placed in a culture dish, and 1.5×104 cells of 786-O were plated in each well and grown to form a confluent and homogeneous layer. Twenty-four hours after cell seeding, the culture insert was removed and a cell-free area, the wound made by the culture insert, could be observed. The cells were incubated with RPMI containing 0.5% FBS, and treated with different concentrations of kaempferol for 0, 6, and 24 h. Cells were photographed using a phase-contrast microscope (×40).
Determination of MMP-2 by zymography
Cells were seeded onto 24-well plates at a density of 3×104 cells/well and treated with kaempferol for 24 h. After indicated treatments, the conditioned media were collected, centrifuged to remove any cellular contaminants and stored at -80℃ until use. Collected media were prepared with sodium dodecyl sulphate (SDS) sample buffer without boiling or reduction and subjected to gelatin zymography analysis to determine the MMPs. Collected media were subjected to 0.1% gelatin-8% SDS polyacrylamide gel electrophoresis to determine the MMPs. The gels were washed with 2.5% Triton X-100 after electrophoresis and incubated in reaction buffer. The gel was then stained with Coomassie brilliant blue R-250.
Western Blot Analysis
786-O cells were seeded onto 10-cm dishes at a density of 6×105 cells and treated with kaempferol at a concentration of 0-100 μM at 37℃ for 24 h. Cell lysates were separated in a 10% polyacrylamide gel and transferred onto a nitrocellulose membrane. The blot was then incubated with standard blocking buffer for 1 h to block nonspecific binding and subsequently incubated with polyclonal antibodies against extracellular signal-regulated protein kinases 1 and 2 (ERK1/2), p-ERK1/2 (Thr185/Tyr187), Akt, p-Akt (Ser473) (Invitrogen Corporation, Carlsbad, CA), focal adhesion kinase (FAK) (Cell Singling Technology, Inc., Danvers, MA) and monoclonal antibodies against p-FAK (Tyr397), phosphatidylinositol 3-kinase (PI3K) (BD Biosciences, Fairleigh, NJ) and MMP-2 (millipore, CA). The signal was detected by enhanced chemiluminescence and quantified using a densitometer (AlphaImager 2000, Alpha Innotech Corporation, San Leandro, CA, USA).
Measurement of Tumor Metastasis in SCID Mice
5-6 weeks old severe combined immunodeficient (SCID) mice (C.B17/IcrPrkdcscid/CrlNarl) weighing 20-23 g were used. The mice were housed with a regular 12-h light/12-h dark cycle and ad libitum access to standard rodent chow diet (Laboratory Rodent Diet 5001, LabDiet, St. Louis, MO), and kept in a pathogen-free environment at the Laboratory Animal Unit. RCC 786-O cells (1×106/0.1 mL/mouse) were injected into SCID mice via their tail veins. After implantation, the mice were randomly divided into three groups (N = 3 for each group) and fed with oral gavage with placebo (saline, control group) and kaempferol (2 and 10 mg/kg/day) suspended in saline. After 150 days, the animals were euthanized with CO2, and the lungs were weighed and examined microscopically with hematoxylin and eosin stain (×200) [24].
Statistical Analysis
One-way analysis of variance (Sigma-Stat 2.0, Jandel Scientific, San Rafael, CA USA) was used to test the statistical significances of the diffenrences throughout this study. A difference with P of <0.05 is considered statistically significant.
Results
Effects of Kaempferol on RCC 786-O Cell Viability
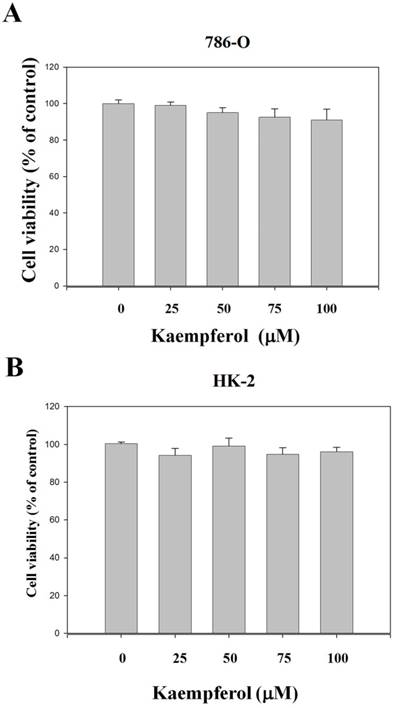
Various concentrations of kaempferol (0, 25, 50, 75, and 100 μM) did not exert significant cytotoxic effects on the RCC 786-O cells. Data from microculture tetrazolium assay (Figure 1A) reveal that the cell numbers of RCC 786-O cells in the presence of 25, 50, 75, and 100 μM kaempferol were not significantly different from those of the control group (0 μM). Using the same procedures, we found that this compound did not exert any significant cytotoxicity on nonmalignant human proximal tubule epithelial HK-2 cells (Figure 1B).
Effects of kaempferol on cell viability of RCC cells. (A) 786-O and (B) HK-2 cells were treated with kaempferol for 24 h by MTT assay. Results were statistically evaluated through one-way ANOVA with post-hoc Dunnett's test. Results from three repeated and separated experiments were similar.
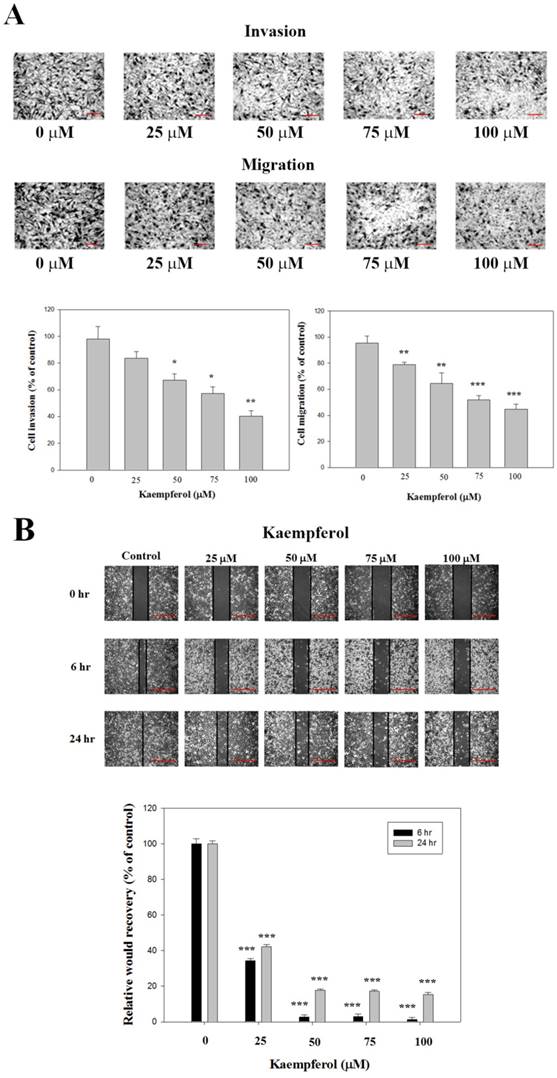
Effects of Kaempferol on RCC 786-O Cell Invasion and Migration
Through cell migration and invasion assay in a 48-well Boyden chamber, the RCC 786-O cells were treated with different concentrations of kaempferol for 24 h. The results show that kaempferol significantly decreased the invasion and migration of RCC 786-O cells (Figure 2A). The results from the wound healing assay show that kaempferol significantly attenuated cell migration dose-dependently in 786-O cells (Figure 2B).
Effects of kaempferol on the cell invasion and migration of RCC cell lines. (A) 786-O cells were treated with kaempferol for 24 h by Boyden chamber invasion and migration assay. Scale bar, 100 μm. (B) The wound healing assay was conducted as described in the Materials and Methods section after 786-O cells were treated with different concentrations of kaempferol for 6 and 24 h. Scale bar, 50 μm. Results were statistically evaluated by using one-way ANOVA with post hoc Dunnett's test (*: P < 0.05; **: P < 0.01; ***: P < 0.001). Results from three repeated and separated experiments were similar.
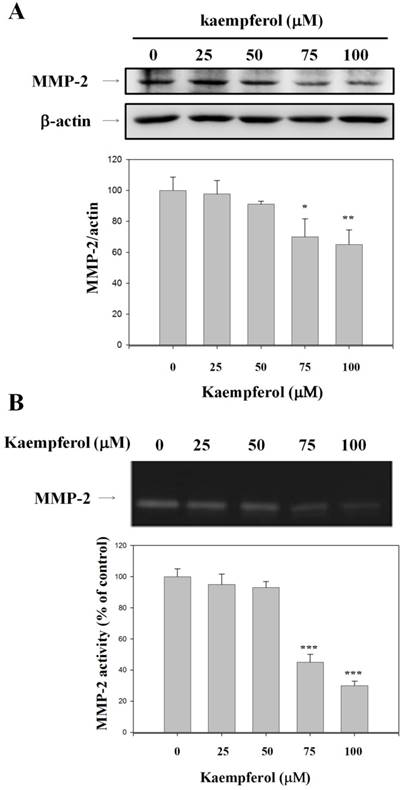
Effects of Kaempferol on the Expression and Activity of MMP-2 in RCC 786-O Cells
To verify the essential role of MMP-2 in cell migration inhibition, we examined MMP-2 protein expression in kaempferol-treated and control RCC 786-O cells. Kaempferol reduced MMP-2 expression, as indicated by the Western blot assay results (Figure 3A). To clarify whether the activities of MMP-2 was involved in inhibiting the invasion of 786-O cells by kaempferol, the effects of kaempferol on MMP-2 activity was investigated by gelatin zymography. Kaempferol reduced the activity of MMP-2 by 69.6% (Figure 3B).
Inhibitory effects of kaempferol on the expression and activity of MMP-2. 786-O cells were treated with kaempferol for 24 h. (A) Western blot analysis of MMP-2 with β-actin as an internal control. (B) MMP-2 activity was analyzed by gelatin zymography. Data represent mean ± SD, with that of control being 100%, and the statistical significance of results was analyzed using one-way ANOVA with post-hoc Dunnett's test (*: P < 0.05; **: P < 0.01).
Effects of Kaempferol on PI3K/Akt and mitogen-activated protein kinase (MAPK) Pathways
For further clarifying the possible underlying mechanisms of inhibition of MMP-2 by kaempferol, the effects of kaempferol on PI3K/Akt and MAPK pathways were examined by Western blot analysis. Results show that a dose-dependent inhibition effect of phosphorylation of Akt in kaempferol-treated RCC 786-O cells (Figure 4A). However, the phosphorylation of ERK1/2 was not significantly inhibited by kaempferol in our results (Figure 4B). Therefore, the inhibition of PI3K/Akt pathway by kaempferol may result in the reduced activity of MMP-2 and tumor invasion.
Effects of kaempferol on the expression levels of PI3K/Akt, ERK1/2, and FAK protein. (A) Western blot analysis of PI3K, p-Akt, and total-Akt, with β-actin as an internal control in 786-O cells after 24 h of treatment with kaempferol. (B) Western blotting with anti-p-FAK, total-FAK, p-ERK1/2, total-ERK1/2 antibodies, with anti-β-actin as an internal control. Similar results were obtained from three repeated and independent experiments. Data represent mean ± SD, with that of control being 100%, and the statistical significance of results was analyzed using one-way ANOVA with post-hoc Dunnett's test (**: P < 0.01; ***: P < 0.001).
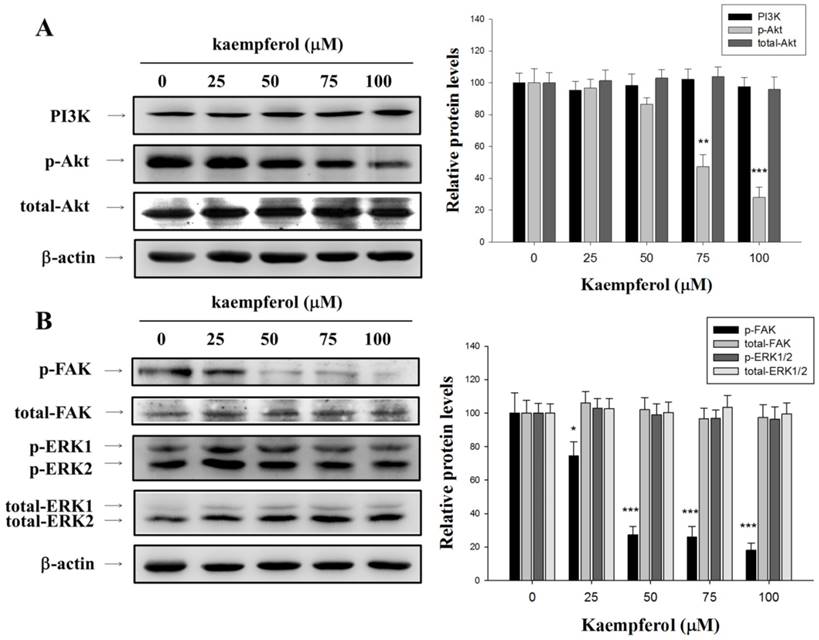
Effects of Kaempferol on FAK Activation
We investigated the molecular regulation of cell migration by kaempferol through Western blot. We then examined the variation in FAK levels in these cells by using antibodies directed against the FAK phosphorylation site Tyr925. As illustrated in Figure 4B, kaempferol reduced FAK phosphorylation. On the basis of these results, we suggested that the inhibition of kaempferol on RCC 786-O cell migration may partly occur through the downregulation of FAK phosphorylation.
Effects of Kaempferol on the Tumor Metastasis of RCC 786-O cells in SCID Mice
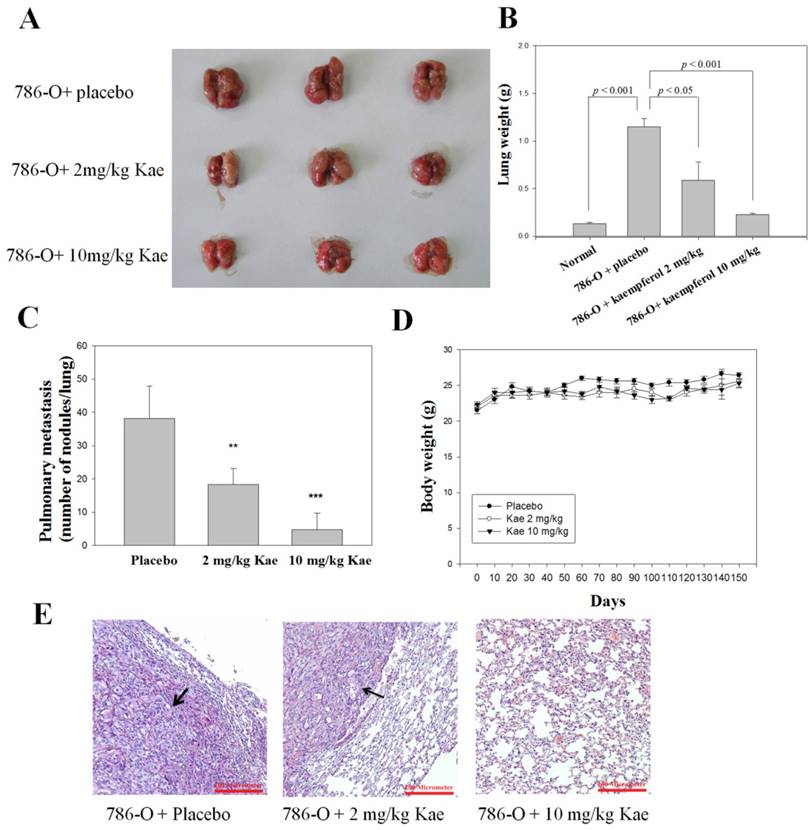
We also performed an in vivo antitumor metastasis study with SCID mice as models by injecting RCC 786-O cells via the tail veins of the mice. Lung tumor nodules were observed especially in the placebo group of mice after 150 days of tumor cell injection (Figure 5A). A significant reduction of lung weight was observed between the placebo and treatment groups (Figure 5B). Vehicle-treated control animals showed massive tumor growth and were given an arbitrary-maximum countable number of about 38.2 ± 9.6. The number was reduced to 4.8 ± 4.9 (10 mg/kg/day; P < 0.001) countable colonies by kaempferol treatment (Figure 5C). Mice body weights were monitored throughout the experiment (Figure 5D). Histopathology of the lungs also showed that the lungs of kaempferol-treated animals had marked reduction in terms of tumor mass (Figure 5E). Accordingly, kaempferol can inhibit the metastasis of RCC 786-O cells in the animal model.
In vivo antimetastatic effects of kaempferol. 786-O cells were injected into the tail veins of six-week-old female SCID mice. After injection of cells, kaempferol (Kae, 2 and 10 mg/kg/d) and placebo (vehicle alone) were administered oral gavage for 150 days. Mice were sacrificed and then analyzed for representative photographs of (A) lungs, (B) lung weight, (C) the number of lung metastasis (D) and body weight in mice. (E) Histopathology of lung of metastatic tumor-bearing animals. Lungs of the metastasis-induced animals were fixed in neutral buffered formalin, and stained with hematoxylin and eosin. Arrows show areas of metastatic nodules (tumor). Results were statistically evaluated by using one-way ANOVA with post-hoc Dunnett's test (**: P < 0.01; ***: P < 0.001).
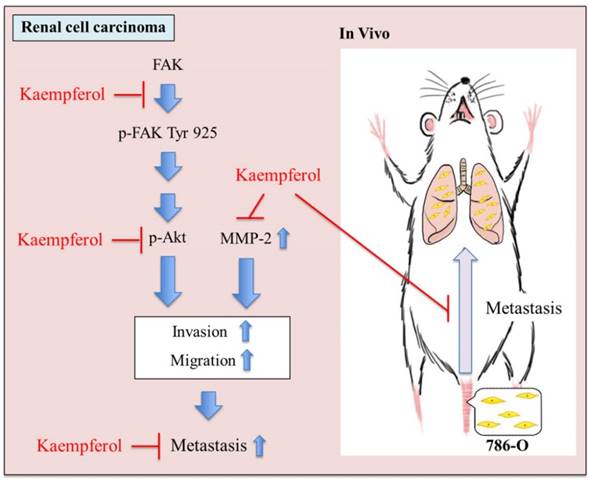
In summary, these findings suggested that kaempferol is able to regulated MMP-2 activity via the down-regulation of FAK-Akt pathways. Finally, kaempferol suppressed cell invasion, migration, and metastasis (Figure 6).
Proposed molecular targets in anti-metastatic efficacy of kaempferol. Kaempferol targets FAK and Akt-mediated expression of MMP-2 and inhibits cell invasion,migration, and metastasis of 786-O cells.
Discussion
A considerable improvement in the 5-year survival rate of patients with renal cell carcinoma has been observed in the last 50 years. However, the long-term survival rate of advanced renal cell carcinoma remains extremely low [5, 25]. Metastatic disease is the leading cause of the poor outcome of advanced renal cell carcinoma. Our study illustrates that kaempferol can inhibit the metastasis of renal cell carcinoma either in the RCC 786-O cell line model or in the animal model.
First, kaempferol inhibits the migration and invasion of RCC 786-O cells. Second, MMP-2 protein expression is suppressed by kaempferol. Third, kaempferol downregulates the phosphorylation levels of Akt and FAK. Finally, the animal model of SCID mice also manifests that kaempferol can inhibit the lung metastasis of the RCC 786-O cell.
Cancer cell metastasis includes several complex processes. Among these processes, only ECM degradation may perform a fundamental role in cancer progress. In particularly, ECM degradation causes metastasis by enabling the spread of cancer cells through the blood and lymphatic vessels [26]. The levels of MMPs, a family of zinc-dependent ECM endopeptidases, are significantly elevated in metastatic human cancers [27]. Carlos reported that hyaluronan promotes cancer cell migration and increases the secretion of matrix metalloproteinase, specifically the increased active form of MMP-2 [28]. Several studies indicated that a high MMP-2 expression is correlated to poor prognosis in prostate cancer [11], bladder cancer, and breast cancer [29]. In addition, MMP-2 upregulation is associated with poor prognosis in RCC patients, and thus the inhibition of MMP-2 activity can suppress cancer metastasis [30]. Moreover, increased MMP-2 expression is more likely to present lymph node metastasis and exhibit rapid progression and poor prognosis [31]. In the study, the inhibition of MMP-2 expression by kaempferol may provide several preventive and therapeutic effects against RCC metastasis. However, the mechanism of MMP-2 upregulation and its signaling pathway involved in the regulation of RCC cell metastasis requires further investigation.
The role of TIMP-2 in metastasis remains ambiguous. Several studies have asserted that increased TIMP-2 levels can protect the hosts from cancer metastasis. However, several researchers have suggested that high TIMP-2 expression results in tumor recurrence. Other researchers reported that the balance between MMP-2 and TIMP-2 can perform an essential part in cancer metastasis. In our study, no significant alteration in TIMP-2 activity was observed (data not shown). Thus, the inhibition of metastasis of the RCC 786-O cells by kaempferol possibly occurred through the suppression of MMP-2 without a change in TIMP-2.
In a recent study, Jo showed that kaempferol suppresses EMT and cell migration in A549 lung cancer cells by inhibiting the Akt1-mediated phosphorylation of Smad3 [32]. Meanwhile, Youyou reported that kaempferol inhibits cholangiocarcinoma growth and metastasis in vitro and in vivo. This finding is related to the inactivation of PI3K/AKT pathway and downstream proteins, which are involved in proliferation, apoptosis, and invasion [33]. In our study, we can see that kaempferol downregulates FAK and Akt phosphorylation levels. However, no significant change was observed in the ERK and JNK/p38 expression levels (data not shown) in our study. Our results demonstrate that suppression of the phosphorylation of FAK and Akt protein and inhibition of the migration of RCC 786-O cell line are consistent with the role of FAK/PI3K/Akt in human dental pulp stem cell migration [34].
FAK, a protein tyrosine kinase, is overexpressed in several cancers and promotes cancer progression and metastasis [35]. The functions of integrins are strongly dependent on FAK activation and its downstream signaling, including the PI3K/Akt-dependent and Ras/MAPK-dependent pathways [36, 37]. Integrin β1/FAK or Akt pathways regulates MMP-2 and MMP-9 expression levels in numerous tumors [38, 39]. Furthermore, FAK protein overexpression has been reported in metastatic human colorectal, breast, thyroid, and prostate cancer cells [40-43]. The knockdown or deletion of FAK impairs tumor growth and the metastatic progression of mammary tumor cells [43]. The PI3K/Akt pathway is aberrantly activated in several types of cancers, and targeting this pathway with drug inhibitors may demonstrate improved anticancer treatment against solid and hematologic tumors [44].
Kaempferol can inhibit RCC 786-O cell migration and invasion partly through MMP-2 suppression, which results from the inhibition of the Akt and FAK pathways but not the MAPK pathways.
In our study, we used RCC 786-O cell and SCID mice to see the suppression effect of kaempferol on RCC 786-O cell lung metastasis. The results show that the lung mass weight in mice treated with 2 mg/kg of kaempferol had a twofold reduction compared with the lung mass weight of the mice in the placebo group (P < 0.05). More than fourfold decline in lung mass weight was observed in group treated with 10 mg/kg of kaempferol (P < 0.001).
In conclusion, the data from our study suggest that kaempferol can inhibit invasion and migration of RCC 786-O cell in vitro by downregulating MMP-2 expression through the inhibition of FAK and Akt phosphorylation levels. In vivo, kaempferol can exert an anti-metastatic effect on the RCC 786-O cells. Therefore, kaempferol may perform a potential role in the prevention and treatment of metastatic RCC.
Acknowledgements
This work was supported by grants from Chung Shan Medical University Hospital [CSH-2015-C-024 and CSH-2012-C-015].
Competing Interests
The authors have declared that no competing interest exists.
References
1. Chow WH, Dong LM, Devesa SS. Epidemiology and risk factors for kidney cancer. Nat Rev Urol. 2010;7:245-257
2. Thompson RH, Ordonez MA, Iasonos A, Secin FP, Guillonneau B, Russo P, Touijer K. Renal cell carcinoma in young and old patients-is there a difference? J Urol. 2008;180:1262-1266 discussion 1266
3. Siemer S, Hack M, Lehmann J, Becker F, Stockle M. Outcome of renal tumors in young adults. J Urol. 2006;175:1240-1243 discussion 1243-1244
4. Cook A, Lorenzo AJ, Salle JL, Bakhshi M, Cartwright LM, Bagi D, Farhat W, Khoury A. Pediatric renal cell carcinoma: single institution 25-year case series and initial experience with partial nephrectomy. J Urol. 2006;175:1456-1460 discussion 1460
5. Pantuck AJ, Zisman A, Belldegrun AS. The changing natural history of renal cell carcinoma. J Urol. 2001;166:1611-1623
6. Stetler-Stevenson WG, Liotta LA, Kleiner DE Jr. Extracellular matrix 6: role of matrix metalloproteinases in tumor invasion and metastasis. FASEB J. 1993;7:1434-1441
7. Stamenkovic I. Matrix metalloproteinases in tumor invasion and metastasis. Semin Cancer Biol. 2000;10:415-433
8. Chien MH, Ying TH, Hsieh YS, Chang YC, Yeh CM, Ko JL, Lee WS, Chang JH, Yang SF. Dioscorea nipponica Makino inhibits migration and invasion of human oral cancer HSC-3 cells by transcriptional inhibition of matrix metalloproteinase-2 through modulation of CREB and AP-1 activity. Food Chem Toxicol. 2012;50:558-566
9. Ranuncolo SM, Armanasco E, Cresta C, Bal De Kier Joffe E, Puricelli L. Plasma MMP-9 (92 kDa-MMP) activity is useful in the follow-up and in the assessment of prognosis in breast cancer patients. Int J Cancer. 2003;106:745-751
10. Bugdayci G, Kaplan T, Sezer S, Turhan T, Koca Y, Kocer B, Yildirim E. Matrix metalloproteinase-9 in broncho-alveolar lavage fluid of patients with non-small cell lung cancer. Exp Oncol. 2006;28:169-171
11. Sauer CG, Kappeler A, Spath M, Kaden JJ, Michel MS, Mayer D, Bleyl U, Grobholz R. Expression and activity of matrix metalloproteinases-2 and -9 in serum, core needle biopsies and tissue specimens of prostate cancer patients. Virchows Arch. 2004;444:518-526
12. Chang HR, Chen PN, Yang SF, Sun YS, Wu SW, Hung TW, Lian JD, Chu SC, Hsieh YS. Silibinin inhibits the invasion and migration of renal carcinoma 786-O cells in vitro, inhibits the growth of xenografts in vivo and enhances chemosensitivity to 5-fluorouracil and paclitaxel. Mol Carcinog. 2011;50:811-823
13. Song N, Sung H, Choi JY, Han S, Jeon S, Song M, Lee Y, Park C, Park SK, Lee KM, Yoo KY, Noh DY, Ahn SH, Lee SA, Kang D. Preoperative serum levels of matrix metalloproteinase-2 (MMP-2) and survival of breast cancer among Korean women. Cancer Epidemiol Biomarkers Prev. 2012;21:1371-1380
14. Ren W, Qiao Z, Wang H, Zhu L, Zhang L. Flavonoids: promising anticancer agents. Med Res Rev. 2003;23:519-534
15. Manthey JA, Grohmann K, Guthrie N. Biological properties of citrus flavonoids pertaining to cancer and inflammation. Curr Med Chem. 2001;8:135-153
16. Liang SQ, Marti TM, Dorn P, Froment L, Hall SR, Berezowska S, Kocher G, Schmid RA, Peng RW. Blocking the epithelial-to-mesenchymal transition pathway abrogates resistance to anti-folate chemotherapy in lung cancer. Cell Death Dis. 2015;6:e1824
17. Li C, Zhao Y, Yang D, Yu Y, Guo H, Zhao Z, Zhang B, Yin X. Inhibitory effects of kaempferol on the invasion of human breast carcinoma cells by downregulating the expression and activity of matrix metalloproteinase-9. Biochem Cell Biol. 2015;93:16-27
18. Lin CW, Chen PN, Chen MK, Yang WE, Tang CH, Yang SF, Hsieh YS. Kaempferol reduces matrix metalloproteinase-2 expression by down-regulating ERK1/2 and the activator protein-1 signaling pathways in oral cancer cells. PLoS One. 2013;8:e80883
19. Labbe D, Provencal M, Lamy S, Boivin D, Gingras D, Beliveau R. The flavonols quercetin, kaempferol, and myricetin inhibit hepatocyte growth factor-induced medulloblastoma cell migration. J Nutr. 2009;139:646-652
20. Lee HS, Cho HJ, Kwon GT, Park JH. Kaempferol Downregulates Insulin-like Growth Factor-I Receptor and ErbB3 Signaling in HT-29 Human Colon Cancer Cells. J Cancer Prev. 2014;19:161-169
21. Kim KY, Jang WY, Lee JY, Jun do Y, Ko JY, Yun YH, Kim YH. Kaempferol Activates G(2)-Checkpoint of the Cell Cycle Resulting in G(2)-Arrest and Mitochondria-Dependent Apoptosis in Human Acute Leukemia Jurkat T Cells. J Microbiol Biotechnol. 2016;26:287-294
22. Marfe G, Tafani M, Indelicato M, Sinibaldi-Salimei P, Reali V, Pucci B, Fini M, Russo MA. Kaempferol induces apoptosis in two different cell lines via Akt inactivation, Bax and SIRT3 activation, and mitochondrial dysfunction. J Cell Biochem. 2009;106:643-650
23. Kang JW, Kim JH, Song K, Kim SH, Yoon JH, Kim KS. Kaempferol and quercetin, components of Ginkgo biloba extract (EGb 761), induce caspase-3-dependent apoptosis in oral cavity cancer cells. Phytother Res. 2010;24(Suppl 1):S77-82
24. Chu SC, Yu CC, Hsu LS, Chen KS, Su MY, Chen PN. Berberine reverses epithelial-to-mesenchymal transition and inhibits metastasis and tumor-induced angiogenesis in human cervical cancer cells. Mol Pharmacol. 2014;86:609-623
25. Hollingsworth JM, Miller DC, Daignault S, Hollenbeck BK. Rising incidence of small renal masses: a need to reassess treatment effect. J Natl Cancer Inst. 2006;98:1331-1334
26. Klein T, Bischoff R. Physiology and pathophysiology of matrix metalloproteases. Amino Acids. 2011;41:271-290
27. Gialeli C, Theocharis AD, Karamanos NK. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011;278:16-27
28. Torre C, Wang SJ, Xia W, Bourguignon LY. Reduction of hyaluronan-CD44-mediated growth, migration, and cisplatin resistance in head and neck cancer due to inhibition of Rho kinase and PI-3 kinase signaling. Arch Otolaryngol Head Neck Surg. 2010;136:493-501
29. Rabi ZA, Todorovic-Rakovic N, Vujasinovic T, Milovanovic J, Nikolic-Vukosavljevic D. Markers of progression and invasion in short term follow up of untreated breast cancer patients. Cancer Biomark. 2015;15:745-754
30. Roy R, Louis G, Loughlin KR, Wiederschain D, Kilroy SM, Lamb CC, Zurakowski D, Moses MA. Tumor-specific urinary matrix metalloproteinase fingerprinting: identification of high molecular weight urinary matrix metalloproteinase species. Clin Cancer Res. 2008;14:6610-6617
31. Cheng HP, Duan YR, Li Y, Li XD, Zhu CY, Chen BP. Clinicopathological Significance of Matrix Metalloproteinase-2 Protein Expression in Renal Cell Carcinoma Patients. Anal Quant Cytopathol Histpathol. 2015;37:353-363
32. Jo E, Park SJ, Choi YS, Jeon WK, Kim BC. Kaempferol Suppresses Transforming Growth Factor-beta1-Induced Epithelial-to-Mesenchymal Transition and Migration of A549 Lung Cancer Cells by Inhibiting Akt1-Mediated Phosphorylation of Smad3 at Threonine-179. Neoplasia. 2015;17:525-537
33. Qin Y, Cui W, Yang X, Tong B. Kaempferol inhibits the growth and metastasis of cholangiocarcinoma in vitro and in vivo. Acta Biochim Biophys Sin (Shanghai). 2016;48:238-245
34. Li M, Sun X, Ma L, Jin L, Zhang W, Xiao M, Yu Q. SDF-1/CXCR4 axis induces human dental pulp stem cell migration through FAK/PI3K/Akt and GSK3beta/beta-catenin pathways. Sci Rep. 2017;7:40161
35. Sulzmaier FJ, Jean C, Schlaepfer DD. FAK in cancer: mechanistic findings and clinical applications. Nat Rev Cancer. 2014;14:598-610
36. Tancioni I, Uryu S, Sulzmaier FJ, Shah NR, Lawson C, Miller NL, Jean C, Chen XL, Ward KK, Schlaepfer DD. FAK Inhibition disrupts a beta5 integrin signaling axis controlling anchorage-independent ovarian carcinoma growth. Mol Cancer Ther. 2014;13:2050-2061
37. Lengyel E. Ovarian cancer development and metastasis. Am J Pathol. 2010;177:1053-1064
38. Gao Y, Guan Z, Chen J, Xie H, Yang Z, Fan J, Wang X, Li L. CXCL5/CXCR2 axis promotes bladder cancer cell migration and invasion by activating PI3K/AKT-induced upregulation of MMP2/MMP9. Int J Oncol. 2015;47:690-700
39. Chen JS, Huang XH, Wang Q, Huang JQ, Zhang LJ, Chen XL, Lei J, Cheng ZX. Sonic hedgehog signaling pathway induces cell migration and invasion through focal adhesion kinase/AKT signaling-mediated activation of matrix metalloproteinase (MMP)-2 and MMP-9 in liver cancer. Carcinogenesis. 2013;34:10-19
40. Tremblay L, Hauck W, Aprikian AG, Begin LR, Chapdelaine A, Chevalier S. Focal adhesion kinase (pp125FAK) expression, activation and association with paxillin and p50CSK in human metastatic prostate carcinoma. Int J Cancer. 1996;68:164-171
41. Owens LV, Xu L, Craven RJ, Dent GA, Weiner TM, Kornberg L, Liu ET, Cance WG. Overexpression of the focal adhesion kinase (p125FAK) in invasive human tumors. Cancer Res. 1995;55:2752-2755
42. Cance WG, Harris JE, Iacocca MV, Roche E, Yang X, Chang J, Simkins S, Xu L. Immunohistochemical analyses of focal adhesion kinase expression in benign and malignant human breast and colon tissues: correlation with preinvasive and invasive phenotypes. Clin Cancer Res. 2000;6:2417-2423
43. Ayaki M, Komatsu K, Mukai M, Murata K, Kameyama M, Ishiguro S, Miyoshi J, Tatsuta M, Nakamura H. Reduced expression of focal adhesion kinase in liver metastases compared with matched primary human colorectal adenocarcinomas. Clin Cancer Res. 2001;7:3106-3112
44. Steelman LS, Stadelman KM, Chappell WH, Horn S, Basecke J, Cervello M, Nicoletti F, Libra M, Stivala F, Martelli AM, McCubrey JA. Akt as a therapeutic target in cancer. Expert Opin Ther Targets. 2008;12:1139-1165
Author contact
![]() Corresponding authors: Horng-Rong Chang MD, PhD, Division of Nephrology, Department of Medicine, Chung Shan Medical University Hospital,Taichung, Taiwan. E-mail: chrcsmucom, TEL: +886-4-24739595 ext. 34704; Yih-Shou Hsieh, Clinical Laboratory, Chung Shan Medical University, Taichung, Taiwan. Email: csmcyshedu.tw, TEL: +886-4-2473-0022
Corresponding authors: Horng-Rong Chang MD, PhD, Division of Nephrology, Department of Medicine, Chung Shan Medical University Hospital,Taichung, Taiwan. E-mail: chrcsmucom, TEL: +886-4-24739595 ext. 34704; Yih-Shou Hsieh, Clinical Laboratory, Chung Shan Medical University, Taichung, Taiwan. Email: csmcyshedu.tw, TEL: +886-4-2473-0022